|
Vernacular Name
Rana de casco de Buckley
State/Provinces
Napo, Orellana, Pastaza, Sucumbíos
Natural Regions
Bosque Piemontano Oriental, Bosque Húmedo Tropical Amazónico
Identification
Description
Osteocephalus buckleyi es una rana arbórea de tamaño moderado que presenta la siguiente combinación de caracteres (modificado de Duellman (7)): (1) el hocico es truncado y la cabeza es casi tan larga como ancha; (2) los dedos de la mano tienen membranas que se extienden alrededor de un tercio de su longitud y en los dedos de los pies alrededor de dos tercios; (3) la piel dorsal en machos lleva una mezcla de tubérculos grandes y pequeños con puntas queratinizadas, mientras que en hembras adultas los tubérculos dorsales son más dispersos; (4) la piel en el vientre es granular, mientras que en el tercio anterior de los flancos es areolada; (5) el tímpano es por lo menos dos tercios del tamaño del ojo.
Color in Life
El dorso es verde pálido con manchas irregulares obscuras; una línea estrecha media dorsal café clara o amarilla puede o no estar presente; los flancos varían de crema a café claro con manchas más obscuras que pueden llegar a ser negras; las superficies anteriores de los muslos son marrones obscuras, y las superficies dorsales y posteriores de los muslos y las pantorrillas son verde apagado, o café claro con manchas verdes o marrones más obscuras; en algunos individuos un tinte azul apagado está presente en las superficie anterior y posterior de los muslos; las membranas son marrones; una amplia barra post-orbital negra se extiende hasta el tímpano, y una amplia barra sub-orbital verde pálida se extiende hasta el labio; el vientre varía de totalmente crema con veteado café obscuro a marrón grisáceo con las puntas de los gránulos blancas mientras que los espacios inter-granulares son marrones; el iris es dorado o bronce verdoso con reticulaciones negras o el color de fondo es uniforme con una línea horizontal marrón; machos en reproducción tienen excrecencias nupciales en las manos de color marrón obscuro (7).
Males Face Cloacal Length Average
Promedio = 41.3 mm (rango 38.0–45.2; n = 14) (18
Females Face Cloacal Length Average
Promedio = 45.7 mm (rango 40.4–50.9; n = 2) (18
Habitat and Biology
En Santa Cecilia (Ecuador), esta especie fue registrada por la noche en vegetación baja en bosque primario o en el borde del bosque (7). En Jatun Sacha y sus alrededores (Ecuador), M. Read (notas del campo) encontró que esta especie es algo común en áreas de bosque secundario y borde de bosque.
Reproduction
Una hembra contenía 580 huevos ováricos maduros (7). El renacuajo es desconocido.
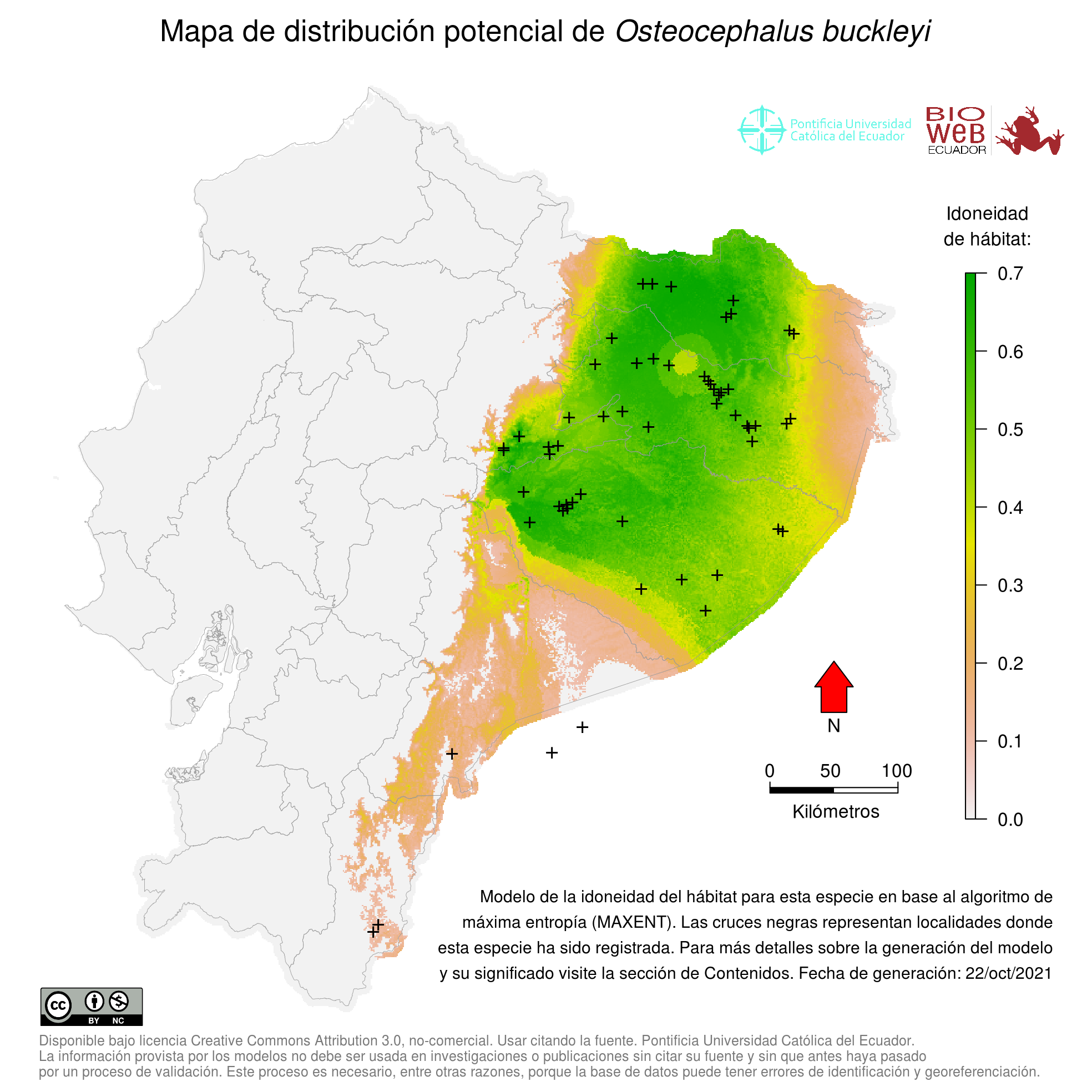
Distribution
Se distribuye en toda la Amazonía de Ecuador y el noreste de Perú (18, 19).
Altitudinal Range
De 0 a 700 m sobre el nivel del mar.
Taxonomy and Evolutionary Relationships
Según la filogenia de Jetz y Pyron (22), su especie hermana es Osteocephalus vilmae. Históricamente, Osteocephalus buckleyi ha sido considerada ser un complejo de varias especies, muchas de ellas crípticas (1, 13). El solapamiento de características morfológicas entre especies condujo a que varias fueran sinonimizadas bajo el nombre Osteocephalus buckleyi por Trueb y Duellman (8); estas especies fueron posteriormente resucitadas (Osteocephalus cabrerai [3], Osteocephalus carri [9], y Osteocephalus festae [13]). La especie con la que más frecuentemente ha sido confundida en la literatura es Osteocephalus cabrerai. A pesar de que Duellman y Mendelson (3) removieron a Osteocephalus cabrerai de la sinonimia con Osteocephalus buckleyi, el espécimen descrito como Osteocephalus cabrerai por Duellman y Mendelson (3) es en realidad un Osteocephalus buckleyi (13). Como resultado de este error, autores subsecuentes (por ejemplo, Gorzula y Señaris (10), Ron y Pramuk (4), Lescure y Marty (11), Lynch (14), Bartlett y Bartlett (12), Vigle (5)) hicieron reportes de Osteocephalus cabrerai que en realidad corresponden a Osteocephalus buckleyi (13, 17).
Ron et al. (1) advirtieron sobre la parafilia encontrada en poblaciones de Osteocephalus buckleyi, las cuales subsecuentemente demostraron ser especies diferentes que fueron descritas por Ron et al. (18). De acuerdo a la filogenia de Ron et al. (18) el grupo de especies O. buckleyi en Ecuador y Perú estaría conformado por cuatro clados con alto soporte que incluyen a 1) Osteocephalus festae + Osteocephalus mutabor, 2) Osteocephalus vilmae + Osteocephalus buckleyi sensu stricto, 3) Osteocephalus verruciger + Osteocephalus cannatellai, y 4) Osteocephalus germani + Osteocephalus cabrerai. Recientemente, Jungfer (19), presentaron una topología algo distinta (en contraste con Ron et al. (18) para su filogenia del grupo buckleyi en la que incluyen especímenes de toda la Amazonía. De acuerdo con este estudio, se incluye también dentro del grupo buckleyi a Osteocephalus mimeticus, Osteocephalus carri y Osteocephalus helenae, así como tres clados que no encajaron filogenéticamente con ninguna otra de las especies del grupo, y posiblemente correspondan a especies nuevas. Todas las especies del grupo buckleyi se reproducen principalmente en riachuelos o zanjas de flujo lento, por lo que esta ha sido propuesta ser una sinapomorfía para el grupo (19).
Jungfer et al. (18) sinonimizaron a Osteocephalus vilmae bajo Osteocephalus buckleyi. Consideramos que Osteocephalus vilmae es una especie válida y que la sinonimia propuesta por Jungfer et al. (18) es incorrecta por dos motivos. Primero, la sinonimia se basa en individuos de dudosa identificación puesto que Jungfer et al. (18) reconocen que no pudieron diferenciar entre ambos grupos genéticos en base a caracteres morfológicos. Sin embargo, en sus comparaciones incluyen individuos no caracterizados genéticamente y que, por lo tanto, solo pudieron ser identificados en base a su morfología. Segundo, Jungfer et al. (18) asumen que eventos de hibridación en condiciones de cautiverio indican la inexistencia de barreras reproductivas en la naturaleza. Este supuesto es incorrecto puesto que la hibridación, con generación de F1 fértiles, es común en cautiverio y en la naturaleza incluso entre especies no hermanas de anfibios (para un ejemplo véase Parris (21). Adicionalmente, la compatibilidad reproductiva con frecuencia es inconsistente con la afinidad evolutiva por lo que asignar poblaciones a una especie en base a ese criterio es lógicamente inconsistente con el estudio del proceso evolutivo (20).
En el mismo trabajo, Jungfer et al. (19) también consideran que Osteocephalus germani, una especie del sur de Perú, es un sinónimo junior de Osteocephalus helenae. Debido a que el holotipo de Osteocephalus helenae es un juvenil y los juveniles de Osteocephalus germani son desconocidos, es imposible, al momento, argumentar que estos binomiales son sinónimos. Por lo tanto, se reconoce a Osteocephalus germani como una especie válida. Se necesitan más estudios para identificar el linaje genético al que pertenece el holotipo de Osteocephalus helenae. Tácitamente, Jungfer et al. (19) asumen la presencia de una sola especie de Osteocephalus del grupo buckleyi en la localidad tipo de Osteocephalus helenae. Sin embargo, la existencia de más de una especie no puede ser descartada sin muestreos exhaustivos.
Etymology
El nombre de la especie hace referencia a Clarence Buckley, el colector del material tipo.
Additional Information
Zimmerman y Bogart (15) proveen una descripción morfológica, características ecológicas de la reproducción, y una descripción detallada del canto de anuncio de individuos de "Osteocephalus buckleyi" de la Amazonía central, cerca de "Manaus", Brasil. Sin embargo, dado nuestro conocimiento actual de la distribución de Osteocephalus buckleyi (presente solo en la Amazonía de Ecuador y noreste de Perú; Jungfer et al. (19)), la información proporcionada por Zimmerman y Bogart (15) probablemente hace referencia a otra especie. Duellman y Mendelson (3) presentan una fotografía y una descripción de un individuo (como Osteocephaus cabrerai). Jungfer (13) muestran una fotografía de una pareja en amplexus. Ron et al. (18) describen el canto, y presentan fotografías a color de una hembra en vista dorso-lateral y ventral, una serie de 10 individuos en vista dorsal y ventral (incluyendo un lectotipo) y de la mano y pie en vista ventral.



Summary Author
Santiago R. Ron, Morley Read y Diego A. Ortiz
Editor
Santiago R. Ron
Edition Date
2013-07-04T00:00:00
Update Date
2022-12-23T16:04:22.38
Literature Cited
Ron, S. R., Toral, E., Venegas, P. J. y Barnes, C. W. 2010. Taxonomic revision and phylogenetic position of Osteocephalus festae (Anura: Hylidae) with description of its larva. Zookeys 70:67–92.PDF Boulenger, G. A.1882. Catalogue of the Batrachia Salientia s. Ecaudata in the collection of the British Museum. Second Edition. London: Taylor & Francis 49530.PDF Duellman, W. E. y Mendelson III, J. R. 1995. Amphibians and reptiles from northern departamento Loreto, Peru: Taxonomy and biogeography. University of Kansas Science Bulletin 55:329-376.PDF Ron, S. R. y Pramuk, J. B. 1999. A new species of Osteocephalus (Anura: Hylidae) from amazonian Ecuador and Peru. Herpetologica 55:433-446.PDF Vigle, G. O. 2008. The amphibians and reptiles of the Estación Biológica Jatun Sacha in the lowland rainforest of Amazonian Ecuador: a 20-year record. Breviora 514:1-27. Rodríguez, L. O. y Duellman, W. E. 1994. Guide to the frogs of the Iquitos Region, Amazonian Perú. Asociación de Ecología y Conservación, Amazon Center for Environmental Education and Research and Natural History Museum, The University of Kansas. Lawrence, Kansas 22:1-80.PDF Duellman, W. E. 1978. The biology of an equatorial herpetofauna in Amazonian Ecuador. Miscellaneous Publications of the University of Kansas 65:1-352.PDF Trueb, L. y Duellman, W. E. 1971. A synopsis of neotropical hylid frogs, genus Osteocephalus. Occasional Papers of the Museum of Natural History, The University of Kansas 1:1-47.PDF Lynch, J. D. 2006. The amphibian fauna in the Villavicencio region of eastern Colombia. La fauna anfibia de la región de Villavicencio en el este de Colombia. Caldasia 28:135-155.PDF Gorzula, S. y Señaris, S. 1998. Contribution to the herpetofauna of the Venezuelan Guayana I. A database. Scientiae Guaianae 8:1-270. Lescure, J. y Marty, C. 2000. Atlas des Amphibiens de Guyane. Collect. Patrimoines Nat., Paris 45:138-139. Bartlett, R. D. y Bartlett, P. P. 2003. Reptiles and Amphibians of the Amazon, An Ecotourist´s Guide. University Press of Florida 1-292. Jungfer, K. 2010. The taxonomic status of some spiny-backed treefrogs, genus Osteocephalus (Amphibia: Anura: Hylidae). Zootaxa 2407:28-50. Lynch, J. D. 2002. A new species of the genus Osteocephalus (Hylidae: Anura) from the western Amazon. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 26:289-292.PDF Zimmerman, B. L. y Bogart, J. P. 1988. Ecology and Calls of Four Species of Amazonian Forest Frogs. Journal of Herpetology 22:97-108. IUCN. 2010. The IUCN red list of threatened species. http://www.iucnredlist.org/apps/redlist/search. (Consultado: 2010). Ron, S. R., Toral, E., Ortiz, D. y Almendáriz, A. 2011. Amphibia, Hylidae, Osteocephalus cabrerai Cochran and Goin, 1970: First confirmed records from Ecuador with distribution map. Check List 7:323–325.PDF Ron, S. R., Venegas, P. J., Toral, E., Read, M., Ortiz, D., Manzano, A. L. 2012. Systematics of the Osteocephalus buckleyi species complex (Anura, Hylidae) from Ecuador and Peru. Zookeys 229:1-52.PDF Jungfer, K., Faivovich, J., Padial, J. M., Castroviejo-Fisher, S., Lyra, M. M., Berneck, B. V. M., Iglesias, P. P., Kok, P., MacCulloch, R. D., Rodrígues, M. T., Verdade, V. K., Torres Gastello, C. P., Chaparro, J. C., Valdujo, P. H., Reichle, S., Moravec, J., Gvozdik, V., Gagliardi-Urrutia, G., Ernst, R., De la Riva, I., Means, D. B., Lima, A. P., Señaris, J. C., Wheeler, W. C., Haddad, C. F. B. 2013. Systematics of spiny-backed treefrogs (Hylidae: Osteocephalus): an Amazonian puzzle. Zoologica Scripta DOI: 10.1111/zsc.12015. Frost, D. R. y Hillis, D. M. 1990. Species in concept and practice: herpetological applications. Herpetologica 46:87-104. Parris, M. J. 1999. Hibridization in leopard frogs (Rana pipiens complex): larval fitness components in single-genotype populations and mixtures. Evolution 53:1872-1883. Jetz, W., Pyron, R. A. 2018. The interplay of past diversification and evolutionary isolation with present imperilment across the amphibian tree of life. Nature Ecology & Evolution 2: 850-858. Ortiz, D., Hoskin, C., Werneck, F., Réjaud, A., Manzi, S., Fouquet, A., Ron, S. 2022. Historical biogeography highlights the role of Miocene landscape changes on the diversification of a clade of Amazonian tree frogs. Organisms Diversity & Evolution 1-20.
|



 Map of Life
Map of Life