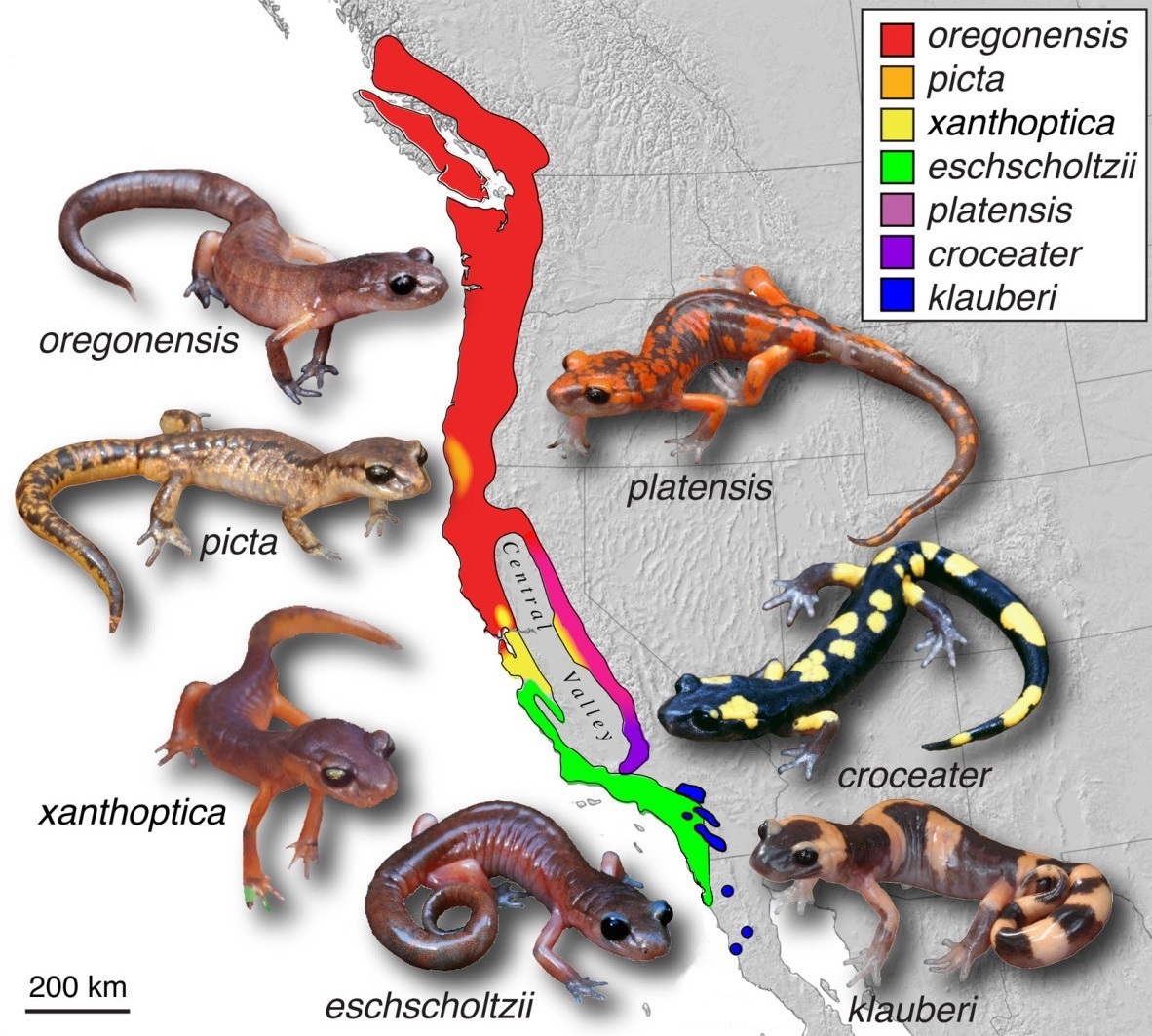
Description Medium-sized, smooth-skinned salamander with 12-13 costal grooves and a distinctive constriction at the base of the swollen tail. Adults range from 45-80 mm snout-vent length to 145 mm total length. Color varies widely and is the basis for subspecies recognition. E. e. oregonensis (Oregon Ensatina) is brown to nearly black above with white to pale yellow below; E. e. eschscholtzii (Monterey Ensatina) is reddish brown above, whitish below; E. e. xanthoptica (Yellow-eyed Ensatina) is orange-brown above, orange below; E. e. picta (Painted Ensatina) is brown above with fine black, yellow, and/or orange blotching, and mottled black and yellow tail; E. e. platensis (Sierra Nevada Ensatina) is gray to brown above, with prominent orange blotching; E. e. croceator (Yellow-blotched Ensatina) is blackish above, with prominent large green-yellow, yellow, or cream blotches; and E. e. klauberi (Large-blotched Ensatina) is blackish above with large orange or pale salmon blotches (all descriptions from Stebbins 1985).
Pacific coast of North America, from extreme SW British Columbia, Canada to northern Baja California, Mexico. Includes Washington, Oregon, and California in the United States. Typical habitat includes coniferous forest, deciduous forest, oak woodland, coastal sage scrub, and chaparral (Stebbins 1954) in thermally buffered mesic microclimates, such as under logs, bark, moss, leaf litter, and talus (Corn and Bury 1991), or in animal burrows (Storer 1925). Notable ring-like distribution of subspecies around the Central Valley of California (Stebbins 1949).
Subspecies ranges are as follows (Stebbins 1985):
E. e. oregonensis, Coast Range and Cascade Mountains from
SW British Columbia to San Francisco Bay;
E. e. eschscholtzii, Coast Range Mountains from San Francisco Bay to NW Baja California; occurs in woodlands in the northern part of the range, but in more open habitats including coastal sage scrub and chaparral in central and southern California;
E. e. xanthoptica, coastal hills east of San Francisco Bay and foothills of central Sierra;
E. e. picta, Coast Range Mountains near Oregon/California border;
E. e. platensis, Sierra of California, from north end of Central Valley to Transverse Ranges;
E. e. croceater, Transverse Ranges at south end of Sierra;
E. e. klauberi, Peninsular Ranges in Southern California.
By Thomas J. Devitt, Stuart J.E. Baird and Craig Moritz (2011) CC BY 2.0, Link
Life History, Abundance, Activity, and Special Behaviors Ensatinas are terrestrial lungless salamanders with direct development (Stebbins 1954; Petranka 1998). Females lay and attend from 3-25 eggs (usually 9-16) in typical aestivation sites at the end of the rainy season (Stebbins 1954; Petranka 1998). Nest sites are often associated with logs (under or within a log, or underneath moss or loose bark on top of the log; summarized in Olson 2006), but may also include rock fissures in seepage areas (Stebbins 1954), animal burrows (Storer 1925) and cavities up to 0.8 m deep (Nussbaum et al. 1983). Juveniles hatch after 114-143 days and appear at the ground's surface at the beginning of winter rains (Stebbins 1954; Petranka 1998).
Ensatinas are usually common where present, with density
estimates of 1730/ha for E. e. xanthoptica (Stebbins 1954) and 2833/ha for E. e. oregonensis (Gneadinger and Reed 1948).
Ensatinas are active during the winter rainy season on very local home ranges, 6-41 meters in width (Stebbins 1954; Staub et al. 1995). During the dry summer, they aestivate in underground retreats such as root cavities and rodent burrows (Stebbins 1954). This species has been shown to mark, recognize, and defend home areas in the laboratory outside the breeding season, suggesting territoriality (Wiltenmuth and Nishikawa 1998).
Coloration in Ensatina shows wide geographic variance, and distinguishes the seven subspecies, as delineated above (see Description). Coloration also serves to deter predators, via crypsis, aposematism (vivid warning colors, associated with toxicity), or mimicry of another species' warning colors. Crypsis via background matching is seen in three Ensatina subspecies: E. e. oregonensis, E. e. picta, and E. e. eschscholtzii (Stebbins 1949); crypsis by way of disruptive coloration is exhibited by another three Ensatina subspecies: E. e. platensis, E. e. croceater, and E. e. klauberi (Stebbins 1949); and Batesian mimicry of aposematic coloration in toxic newts (genus Taricha) is used by E. e. xanthoptica (Kuchta et al. 2008). Subspecies with disruptive coloration (blotching or spotting) are associated with forested areas, usually closed-canopy forest (Stebbins 1949).
Stebbins (1949) proposed that the vivid orange and yellow coloration of E. e. xanthoptica might be mimicry of the highly toxic newts Taricha torosa and Taricha granulosa, both of which are sympatric with E. e. xanthoptica. Kuchta et al. (2005) first tested this hypothesis, and showed that painted clay model salamanders with coloration matching that of E. e. xanthoptica were attacked significantly less often than brown morphs, despite both being equally visible on a white paper base. In experimental trials, Western Scrub-Jays presented first with a newt (Taricha torosa) were significantly less quick to attack and consume the presumed newt mimic E. e. xanthoptica than to attack and consume the cryptically colored E. e. oregonensis (Kuchta et al. 2008). Since jays were observed to eat E. e. xanthoptica tail and all, with relish and without ill effect, E. e. xanthoptica is apparently edible, making its resemblance to the highly toxic Taricha a case of Batesian mimicry rather than Müllerian (Kuchta et al. 2008).
Ensatinas lack traditional amphibian mucous glands as well as lacking typical, acidic mucus, instead secreting neutral mucus via specialized, derived granular glands (Fontana et al. 2006). The toxicity of E. e. xanthoptica has not been evaluated; high concentration of granular glands are found in Ensatina tails, and secretion under stress of a milky, sticky glutinous substance with an astringent taste has been observed (Kuchta et al. 2008). However, since Western Scrub-Jays are able to eat Ensatinas without any apparent ill effect, these salamanders appear not to be particularly toxic (Kuchta et al. 2008). The birds did spend considerable time scraping their beaks during and after consumption of E. e. xanthoptica (Kuchta et al. 2008). The stress-induced, sticky white mucous secretions may actually be an antipredator defense by virtue of being extremely adhesive, enabling Ensatinas to avoid being consumed by predators such as Western Terrestrial garter snakes (Thamnophis elegans; Arnold 1982).
Trends and Threats Probably because Ensatina has its highest abundance in edge habitats (Stebbins 1951), it remains fairly common
throughout most of its range despite human development. Surveys in northwestern California forests, for example, show no evidence of population declines over the time period 1984-1995 (Welsh et al. 2007). The subspecies E. e. croceater and E. e. klauberi, however, inhabit more fragmented habitat in the drier part of Southern California, and are presumably more susceptible to localized extinction. Few threats are known for this species beyond habitat destruction.
Interestingly, while Storer (1925) found that E. e. xanthoptica was less common in central California than Aneides lugubris, the reverse is now true, presumably because Aneides is now less
common (D. B. Wake, pers. comm.)
Direct development away from streams and the dispersed distribution of Ensatinas in a variety of common
habitats likely make it less sensitive than other species with breeding aggregations or migrations or more specific aestivation or brooding sites. However, Olson et al. (2006) found that Ensatina nests in a managed forest in Oregon were preferentially associated (13 of 14) with downed wood, particularly large logs; they concluded that management of downed wood to provide cover and nesting sites may be necessary in young stands of forest. In addition, captures of adult Ensatinas decreased by 40% in thinned forests lacking downed wood, whereas captures in thinned forests remained the same where downed wood volume was high (Rundio and Olson 2007).
Relation to Humans Ensatinas are not commonly part of the pet trade, nor are they hunted or otherwise used by humans. They are often found on the edge of rural human development, however, and artificial debris from those developments (discarded boards, tires, etc.) is often used for cover in those habitats (D. Parks, pers. obs).
Possible reasons for amphibian decline
General habitat alteration and loss Habitat modification from deforestation, or logging related activities Intensified agriculture or grazing Urbanization
Comments The Ensatina complex has long been studied as a textbook case of speciation (Futuyma 1998). The complex exhibits a ring-like distribution around the inhospitable Central Valley of California, with intergradation at the north end of the Valley but sympatry at the southern meeting of Sierran and coastal subspecies (Stebbins 1949). This pattern has inspired numerous studies of genetic diversity within the complex (Wake and Yanev 1986; reviewed in Wake 2006).
This species was featured as News of the Week on 15 May 2017:
Resident microbial communities living on amphibian skin can have significant effects on host health, yet the basic ecology of the host-microbiome relationship of many amphibian taxa is poorly understood. Prado-Irwin et al (2017) used NextGeneration sequencing to characterize intraspecific variation in the skin microbiome of the salamander Ensatina eschscholtzii xanthoptica, a subspecies composed of four genetically distinct populations distributed throughout the San Francisco Bay Area and the Sierra Nevada mountains in California, USA. They found that salamanders from four geographically and genetically isolated populations harbor similar skin microbial communities, and that they were significantly different from those of the surrounding terrestrial environment. These results suggest that the relationship between E. e. xanthoptica salamanders and their resident skin microbiomes is conserved, possibly indicating a stable mutualism between the host and microbiome (Written by Vance Vredenburg).
Watch KQED's Deep Look on the ring species Ensatina
Arnold, S. J. (1982). ''A quantitative approach to antipredator performance: salamander defense against snake attack.'' Copeia, 1982, 247-253.
Corn, P. S. and Bury, R. B. (1991). ''Terrestrial amphibian communities in the Oregon Coast Range.'' Wildlife and Vegetation of Unmanaged Douglas-fir Forests. L. F. Ruggiero, K. B. Aubry, A. B. Carey, and M. H. Huff, eds., USDA Forest Service General Technical Report PNW-GTR-285, 304-317.
Devitt TJ, Baird SJE, Moritz C (2011). "Asymmetric reproductive isolation between terminal forms of the salamander ring species Ensatina eschscholtzii revealed by fine-scale genetic analysis of a hybrid zone." BMC Evolutionary Biology, 11(1), 245-. [link]
Fontana, M. F., Ask, K. A., Macdonald, R. J., Carnes, A. M., and Staub, N. L. (2005). ''Loss of traditional mucus glands and presence of a novel mucus-producing granular gland in the plethodontid salamander Ensatina eschscholtzii.'' Biological Journal of the Linnean Society, 87(3), 469-477.
Gneadinger, L. M., and Reed, C. A. (1948). ''Contribution to the natural history of the plethodontid salamander Ensatina eschscholtzii.'' Copeia, 1948, 187-196.
Kuchta, S. R. (2005). ''Experimental support for aposematic coloration in the salamander Ensatina eschscholtzii xanthoptica: implications for mimicry of Pacific newts.'' Copeia, 2005(2), 265-271.
Kuchta, S. R., Krakauer, A. H., and Sinervo, B. (2008). ''Why does the Yellow-eyed Ensatina have yellow eyes? Batesian mimicry of Pacific Newts (genus Taricha) by the salamander Ensatina eschscholtzii xanthoptica.'' Evolution, 62(4), 984-990.
Nussbaum, R. A., Brodie, E. D., Jr., and Storm, R. M. (1983). Amphibians and Reptiles of the Pacific Northwest. University of Idaho Press, Moscow, Idaho.
Olson, D. H., Naumann, R. S., Ellenburg, L. L., Hansen, B. P., and Chan, S. S. (2006). ''Ensatina eschscholtzii nests at a managed forest site in Oregon.'' Northwestern Naturalist, 87, 203-208.
Petranka, J. W. (1998). Salamanders of the United States and Canada. Smithsonian Institution Press, Washington D.C. and London.
Rundio, D. E., and Olson, D. H. (2007). ''Influence of headwater site conditions and riparian buffers on terrestrial salamander response to forest thinning.'' Forest Science, 53(2), 320-330.
Staub, N. L., Brown, C. W., and Wake, D. B. (1995). ''Patterns of growth and movements in a population of Ensatina eschscholtzii platensis (Caudata: Plethodontidae) in the Sierra Nevada, California.'' Journal of Herpetology, 29, 593-599.
Stebbins, R. C. (1949). "Speciation in salamanders of the Plethodontid genus Ensatina." University of California Publications in Zoology, 48, 377-526.
Stebbins, R. C. (1954). "Natural history of the salamanders of the Plethodontid genus Ensatina." University of California Publications in Zoology, 54(2), 47-124.
Stebbins, R. C. (1985). A Field Guide to Western Reptiles and Amphibians. Houghton Mifflin, Boston.
Stebbins, R.C. (1951). Amphibians of Western North America. University of California Press, Berkeley.
Storer, T. I. (1925). "A synopsis of the amphibia of California." University of California Publications in Zoology, 27, 1-342.
Wake, D. B. (2006). ''Problems with species: patterns and processes of species formation in salamanders.'' Annals of the Missouri Botanical Gardens, 93, 8-23.
Wake, D. B. and Yanev, K. P. (1986). ''Geographic variation in allozymes in a 'ring species,' the plethodontid salamander Ensatina eschscholtzii of western North America.'' Evolution, 40(4), 702-715.
Welsh, H. H., Jr., Fellers, G. M., and Lind, A. J. (2007). ''Amphibian populations in the terrestrial environment: is there evidence of declines of terrestrial forest amphibians in northwestern California?'' Journal of Herpetology, 41(3), 469-482.
Wiltenmuth, E. B. and Nishikawa, K. C. (1998). ''Geographical variation in agonistic behaviour in a ring species of salamander, Ensatina eschscholtzii.'' Animal Behavior, 55, 1595-1606.
Originally submitted by: Duncan Parks (first posted 1999-02-23)
Species Account Citation: AmphibiaWeb 2024 Ensatina eschscholtzii: Ensatina Salamander <https://amphibiaweb.org/species/4046> University of California, Berkeley, CA, USA. Accessed May 11, 2025.



 Raffaëlli Account
Raffaëlli Account Map of Life
Map of Life