|
Vernacular Name
Rana terrestre de André
State/Provinces
Morona Santiago, Napo, Orellana, Pastaza, Sucumbíos, Zamora Chinchipe
Natural Regions
Bosque Húmedo Tropical Amazónico
Identification
Es una rana pequeña de color café o gris con marcas oscuras, flancos con puntos redondeados oscuros y vientre blanco. Tiene el hocico largo y puntiagudo, los dedos I y II de la mano de igual tamaño, sin rebordes cutáneos en los dedos y discos expandidos ausentes. Estas características la diferencian de individuos pequeños de Leptodactylus discodactylus, Leptodactylus leptodactyloides, Leptodactylus petersii y Leptodactylus wagneri que tienen el Dedo I más grande que el Dedo II en la mano, presencia de rebordes cutáneos en los dedos de pies y vientre o garganta con moteado obscuro. El resto de Leptodactylus en la región que no tienen rebordes cutáneos en los dedos de pies, son más grandes (LRC > 29 mm) y tienen pliegues dorsolaterales conspicuos. No obstante, líneas dorsolaterales formadas por tubérculos glandulares son discernibles en Adenomera andreae. Adenomera andreae se asemeja más a Adenomera hylaedactyla, pero se diferencia por poseer un hocico equivalente a una vez el diámetro del ojo (una vez y media el diámetro del ojo en Adenomera hylaedactyla), líneas dorsolaterales glandulares poco visibles (conspicuas en Adenomera hylaedactyla), pecho y superficie ventral de brazos más obscura que el vientre (pecho, superficie ventral de brazos y vientre claros en Adenomera hylaedactyla), y superficie posterior de vientre y superficie anterior de muslos grisáceas (amarillentas en Adenomera hylaedactyla) (20). Usando un análisis de función discriminante, Heyer (15) indica también que Adenomera andreae y Adenomera hylaedactyla son morfológicamente distintas. La ausencia de discos expandios en los dígitos distingue a Adenomera andreae de Pristimantis, y la ausencia de membrana en los dedos de pies la distingue de Scinax y Rana (4, 6). Las poblaciones de Adenomera en Ecuador han sido adscritas a Adenomera andreae o Adenomera hylaedactyla; sin embargo, el estudio de Fouquet et al. (29) indica que ninguna de las dos especies se encontraría en el país (ver Taxonomía y relaciones evolutivas).
Description
Especie de tamaño muy pequeño a pequeño con la siguiente combinación de caracteres (4, 6, 20): (1) hocico moderadamente largo y puntiagudo; distancia entre ojo y punta de hocico es igual al diámetro del ojo; tímpano distinto; (2) dos líneas dorsolaterales delgadas formadas por tubérculos glandulares a cada lado del cuerpo son apenas discernibles; (3) longitud de Dedo I = Dedo II en la mano; almohadillas y espinas nupciales ausentes; (4) dedos de pies carecen de membrana y rebordes cutáneos; (5) dígitos de manos y pies terminan en apenas expandidas y redondeadas en vista dorsal, aplanadas en vista lateral.
Color in Life
Dorso varía de café oliva apagado a café grisáceo con marcas más obscuras, con o sin una línea dorsolateral crema a rosa; una línea media-dorsal crema puede estar presente en la parte posterior del cuerpo; flancos con puntos redondeados obscuros; marca interorbital triangular obscura con el ápice dirigido posteriormente; labio superior con dos barras suborbitales cafés obscuras; superficie dorsal de extremidades posteriores con barras transversales obscuras; superficie posterior de muslos café; vientre blanco con manchas cafés periféricas; región gular, pecho, superficie inferior de brazos y piernas rosadas a grisáceas; iris bronce apagado con reticulaciones o manchas diminutas obscuras (4, 6, 20).
Males Face Cloacal Length Average
Promedio = 23.3 mm. máximo 27.0 (rango 17–20) (5, 20
Females Face Cloacal Length Average
Promedio = 24.1 mm.máximo 28.9 (rango 20–22) (5, 20
Habitat and Biology
Especie terrestre y principalmente diurna, aunque también ha sido observada en la noche; habita en la hojarasca y vegetación baja en bosque primario y secundario (6, 4, 7, 20, 25, 34). Morley Read (notas de campo) reporta haber encontrado a la especie también en borde de bosque en Ecuador. Aunque presentan esencialmente la misma distribución geográfica, Adenomera andreae difiere ecológicamente de Adenomera hylaedactyla en que esta última es nocturna y habita en áreas abiertas (4, 15, 34). Ribeiro et al. (27) encontraron que Adenomera aff. andreae fue encontrada igualmente en sitios riparios como no riparios. Menin et al. (25) indican que especies no dependientes del agua para su reproducción (incluyendo Adenomera aff. andreae) son generalistas y muestran poca relación en cuanto a las características del hábitat a mesoescala. Sin embargo, encontraron que la abundancia de Adenomera aff. andreae disminuye conforme aumenta la pendiente del terreno (25). Adenomera andreae se reproduce principalmente en época lluviosa (7, 9, 20). Los machos vocalizan sobre el suelo desde abajo de la hojarasca y troncos caídos; el canto consiste en una nota larga, sencilla y estridente; el macho escarba un agujero en el suelo donde la hembra es guiada para depositar un nido de espuma que contiene < 20 huevos, de 2–3 mm de diámetro; los renacuajos son endotróficos y se desarrollan completamente dentro de los nidos en los agujeros terrestres (4, 5, 7, 20, 32).
Moreira y Lima (14) reportan que la oviposición de huevos ocurre antes de la estación lluviosa, mientras que el reclutamiento de juveniles durante la estación lluviosa; los juveniles alcanzan el tamaño adulto en 6–8 meses. Kokobum y Giaretta (32) reportan que en una especie (Adenomera sp.) del Cerrado de Brasil, existen interacciones territoriales entre machos y los renacuajos comienzan a generar espuma para el nido transcurridas 10 horas después de la oviposición, lo cual evita la compactación de los mismos y podría incrementar su eficiencia respiratoria y excretoria. Tales aspectos aún requieren ser investigados en Adenomera andreae. Adenomera andreae es una especie generalista en cuanto a su dieta, alimentándose de hormigas, cucarachas, escarabajos, ácaros, colémbolas, termitas, grillos, arañas, cienpiés, milpiés, chinches, cigarras, tijeretas y anélidos (6, 13, 20, 28). Lima y Magnusson (35) encontraron que individuos de Adenomera andreae comen proporciones grandes de ácaros cuando son juveniles y proporciones grandes de hormigas cuando son adultos. Predadores de Adenomera andreae incluyen arañas (Ancylometes rufus y Ctenus spp.) en la Amazonía central (10). Menin y Giaretta (31) mencionan que larvas de moscas (Beckeriella niger) infestan y depredan nidos de leptodactílidos sintópicos como Physaleamus y Leptodactylus, pero no de una especie de Adenomera, probablemente por el diferente grado de exposición al medio ambiente (nidos enterrados en Adenomera).
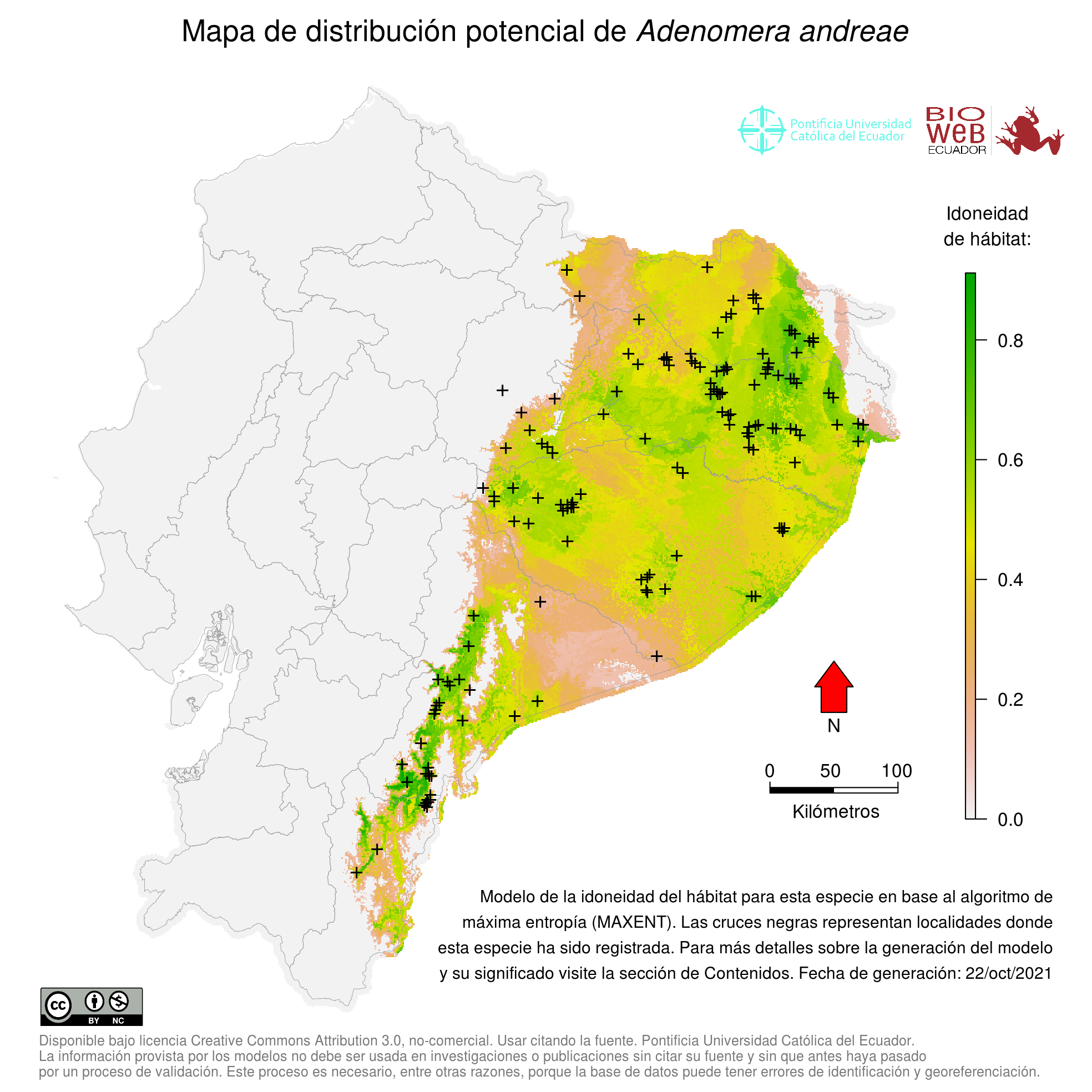
Distribution
Se distribuye en las tierras bajas de la Amazonía al este de los Andes en Brasil, Guyana, Surinam, Guayana Francesa, Venezuela, Colombia, Ecuador y Perú (20, 21, 23, 34). Por otra parte, Fouquet et al. (29) reporta que Adenomera andreae no se encontraría en Ecuador (ver Taxonomía y relaciones evolutivas).
Altitudinal Range
De 0 a 400 m sobre el nivel del mar (21).
Taxonomy and Evolutionary Relationships
Frost (23) provee sinónimos y comentarios taxonómicos. Heyer (5) la ubicó en el grupo de especies Leptodactylus marmoratus, pero posteriormente (11) resucitó el género Adenomera para los miembros de este grupo. Adenomera es un grupo taxonómicamente complejo y los especimenes preservados son de difícil identificación debido a la gran variación morfológica con-interespecífica, varias especies viven en simpatría e incluso sintopía, y existe una gran proporción de especies crípticas que todavía esperan descripción (19, 29). Ciertos autores han presentado evidencia morfológica (5), molecular (23, 26) y reproductiva (32) mostrando a Leptodactylus parafilético con respecto a Adenomera, y consecuentemente han sugerido sinonimizar Adenomera bajo Leptodactylus. Sin embargo, otras líneas de evidencia soportan la distinción de Adenomera como un género diferente de Leptodactylus, y que Lithodytes lineatus es su taxón hermano; a su vez ambos géneros formarían el clado hermano de Leptodactylus. Estas relaciones son soportadas por cariotipos (11), miología y osteología (17), cantos (19), ecología (22), y morfología y filogenética (3, 18, 24, 29, 34). Heyer (22), Kokobum y Giaretta (32), y de Sá et al. (18) reportan que Adenomera comparte características reproductivas con el grupo Leptodactylus fuscus lo que ha llevado a pensar una aparente relación; sin embargo, estas afinidades han evolucionado independientemente y así estos grupos no se encuentran cercanamente relacionados. Heyer (15) presenta un análisis de función discriminante dentro de Adenomera andreae, indicando que no existen diferencias morfológicas significativas en cuanto a variación geográfica. Fouquet et al. (33) reportan que los ríos constituyen barreras efectivas a la dispersión de esta especie.
Fouquet et al. (29) realizaron la revisión más reciente del género Adenomera en base a información molecular. Su estudio reveló que Adenomera se originó en la Amazonía, con subsecuentes dispersiones durante el Mioceno hacia el Diagonal Seco (Chaco, Cerrado y Caatinga) y al Bosque atlántico, las cuales estuvieron asociadas con cambios del bosque hacia hábitats abiertos. Su estudio indica que Adenomera andreae y Adenomera hylaedactyla no se encontrarían en Ecuador, y que el taxón presente en Ecuador es Adenomera simonstuarti, una especie descrita del Cuzco amazónico, Perú y que se encuentra muy relacionada con la nominal Adenomera andreae. Fouquet et al. (29) reconoce que Adenomera simonstuarti estaría conformada por varios subclados con alto soporte (incluyendo aquí muestras de Ecuador), lo que es también corroborado por diferencias en el canto de anuncio de especímenes de Ecuador vs. especímenes de la localidad tipo de Adenomera simonstuarti, revelando así la existencia de más de una especie (3 especies candidatas no confirmadas para Ecuador). Sin embargo, en ausencia de más evidencia, Fouquet et al. (29) conservativamente ubican a estos subclados bajo Adenomera simonstuarti. Anfibios del Ecuador reconoce que Adenomera andreae y Adenomera hylaedactyla probablemente no se encuentren en el país, pero no comparte la decisión de Fouquet et al. (29) en designar a todas las especies de Ecuador bajo Adenomera simonstuarti. Por lo tanto, hasta que un estudio integrativo del género con un enfoque en especímenes de Ecuador pueda ser alcanzado, previsoramente se reconoce a Adenomera andreae y Adenomera hylaedactyla como especies presentes en el país. Por otro lado, en la filogenia de Jetz y Pyron (36) proponen a Adenomera andreae como la especie hermana de un clado conformado por el resto de especies del género.
Additional Information
Pearman (8) y Vigle (2) se refieren a la especie que habita en Jatún Sacha (Provincia de Napo, Ecuador) como Adenomera andreae; sin embargo, la especie de Jatún Sacha es nocturna y se encuentra en bosque secundario, por lo que probablemente corresponda en realidad a Adenomera hylaedactyla. Duellman (6) y Rodríguez y Duellman (4) presentan una fotografía en vista dorsolateral. Bogart (12) describe el cariotipo como 2N = 26 (1 par de cromosomas metacéntricos, 4 submetacéntricos, 2 subtelocéntricos y 6 telocéntricos). Menin et al. (10) presentan fotografías de arañas alimentándose de Adenomera andreae. Lima et al. (20) presentan fotografías de un macho y una hembra en vista dorsolateral, un macho en vista ventral, variación del patrón dorsal y una puesta de huevos. Ponssa y Heyer (16) describen el esqueleto óseo en Adenomera, y sugieren que los dos especímenes analizados de Ecuador (ambos identificados como Adenomera andreae) corresponderían a dos especies distintas. Sin embargo, en ausencia de más líneas de evidencia, no pudieron asignar ninguna de estos especies a Adenomera andreae o Adenomera hylaedactyla.



Summary Author
Diego A. Ortiz, Morley Read y Santiago R. Ron
Editor
Santiago R. Ron
Edition Date
2014-02-07T00:00:00
Update Date
2022-09-26T18:26:29.867
Literature Cited
Müller, L. 1923. Neue order seltene reptilen und batarachier der Zoologischen Sammlung des bayrisehen staates. Zoologischer Anzeiger 57:39-54.PDF Vigle, G. O. 2008. The amphibians and reptiles of the Estación Biológica Jatun Sacha in the lowland rainforest of Amazonian Ecuador: a 20-year record. Breviora 514:1-27. Frost, D. R., Grant, T., Faivovich, J., Bain, R., Günther, A. C., Haas, A., Haddad, C. F., de Sá, R., Donnellan, S. C., Raxworthy, C. J., Wilkinson, M., Channing, A., Campbell, J. A., Blotto, B. L., Moler, P., Drewes, R. C., Nussbaum, R. A., Lynch, J. D., Green, D. y Wheeler, W. C. 2006. The amphibian tree of life. Bulletin of the American Museum of Natural History 297:370.PDF Rodríguez, L. O. y Duellman, W. E. 1994. Guide to the frogs of the Iquitos Region, Amazonian Perú. Asociación de Ecología y Conservación, Amazon Center for Environmental Education and Research and Natural History Museum, The University of Kansas. Lawrence, Kansas 22:1-80.PDF Heyer, W. R. 1973. Systematics of the Marmoratus group of the frog genus Leptodactylus (Amphibia, Leptodactyluidae). Natural History Museum of Los Angeles County Contributions in Science :1-50. Duellman, W. E. 1978. The biology of an equatorial herpetofauna in Amazonian Ecuador. Miscellaneous Publications of the University of Kansas 65:1-352.PDF Toft, C. A. y Duellman, W. E. 1979. Anurans of the lower Río Llullapichis, Amazonian Peru: a preliminary analysis of community structure. Herpetologica 35:71-77.PDF Pearman, P. B. 1997. Correlates of amphibian diversity in an altered landscape of Amazonian Ecuador. Conservation biology 11:1211-1225. Duellman, W. E. 1995. Temporal Fluctuations in Abundances of Anuran Amphibians in a Seasonal Amazonian Rainforest. Journal of Herpetology 29:13-21. Menin, M., Rodrigues, J., de Azevedo, C. S. 2005. Predation on amphibians by spiders (Arachnida, Araneae) in the Neotropical region. Phyllomedusa 41:39-47. Heyer, W. R. 1974. Relationships of the marmoratus species group (Amphibia, Leptodactylidae) within the subfamily Leptodactylinae. Contributions in Science. Natural History Museum of Los Angeles County 253:1-46.PDF Bogart, J. P. 1974. A Karyosystematic Study of Frogs in the genus Leptodactylus (Anura: Leptodactylidae). Copeia 1974:728-737.PDF Toft, C. A. 1980. Feeding Ecology of Thirteen Syntopic Species of Anurans in a Seasonal Tropical Environment. Oecologia 45:131-141.PDF Moreira, G. J. y Lima, A. P. 1991. Seasonal Patterns of Juvenile Recruitment and Reproduction in Four Species of Leaf Litter Frogs in Central Amazonia. Herpetologica 47:295-300. Heyer, W. R. 1977. A discriminant function analysis of the frogs of the genus Adenomera (Amphibia:Leptodactylidae). Proceedings of the Biological Society of Washington 89:581-592. Ponssa, M. L. y Heyer, W. R. 2007. Osteological characterization of four putative species of the genus Adenomera (Anura: Leptodactylidae), with comments on intra- and interspecific variation. Zootaxa 1403:37-54.PDF Heyer, W. R. 1975. A preliminary analysis of the intergeneric relationships of the frog family Leptodactylidae. Smithsonian Contributions to Zoology 199:1-55. De Sá, R. O., Heyer, W. R., Camargo, A. 2005. A Phylogenetic Analysis of Vanzolinius Heyer, 1974 (Amphibia, Anura, Leptodactylidae): Taxonomic and life history implications. Arquivos do Museu Nacional, Río de Janeiro 63:707-726.PDF Angulo, A., Cocroft, R. B. y Reichle, S. 2003. Species identity in the genus Adenomera (Anura: Leptodactylidae) in southeastern Peru. Herpetologica 59:490-504.PDF Lima, A. P., Magnusson, W. E., Menin, M., Erdtmann, L., Rodrigues, D. J., Keller, C., Hödl, W. 2006. Guia de sapos da Reserva Adolpho Ducke, Amazônia Central = Guide to the frogs to Reserva Adolpho Ducke, Central Amazonia. Átemma, Manaus. 168 p. IUCN. 2010. The IUCN red list of threatened species. http://www.iucnredlist.org/apps/redlist/search. (Consultado: 2010). Heyer, W. R. 1969. Studies on the genus Leptodactylus (Amphibia, Leptodactylidae) III. A redefinition of the genus Leptodactylus and a description of a new genus of leptodactylid frogs. Contributions in Science. Natural History Museum of Los Angeles County 155: 1-14.PDF Frost, D. R. 2013. Amphibian Species of the World: an online reference. Version 5.6 (15 October, 2012). Base de datos accesible en http://research.amnh.org/vz/herpetology/amphibia/ American Museum of Natural History, New York, USA. Pyron, A. y Wiens, J. J. 2013. Large-scale phylogenetic analyses reveal the causes of high tropical amphibian diversity. Proceedings of the Royal Society B 280:20131622. Menin, M., Lima, A. P., Magnusson, W. E., Waldez, F. 2007. Topographic and edaphic effects on the distribution of terrestrially reproducing anurans in Central Amazonia: mesoscale spatial patterns. journal of Tropical Ecology 23:539-547. Jansen, M., Bloch, R., Schulze, A., Pfenninger, M. 2011. Integrative inventory of Bolivia’s lowland anurans reveals hidden diversity. Zoologica Scripta 40:567-583. Ribeiro, J. W., Lima, A. P., Magnusson, W. E. 2012. The effect of riparian zones on species diversity of frogs in Amazonian forests. Copeia 2012:375-381. Simon, M. P. y Toft, C. A. 1991. Diet specialization in small vertebrates: Mite-eating in frogs. Oikos 61:263-278. Fouquet, A., Santana Cassini, C., Haddad, C. F. B., Pech, N., Rodrigues, M. T. 2013. Species delimitation, patterns of diversification and historical biogeography of the Neotropical frog genus Adenomera (Anura, Leptodactylidae). Journal of Biogeography. doi: 10.1111/jbi.12250 Dixon, J. R. y Kofron, C. 1983. The Central and South American anomalepid snakes of the genus Liotyphlops. Amphibia-Reptilia 4:241-264. Menin, M. y Giaretta, A. A. 2003. Predation on foam nests of leptodactyline frogs (Anura: Leptodactylidae) by larvae of Beckeriella niger(Diptera: Ephydridae). Journal of Zoology 261:239-243. Kokobum, M. N. C. y Giaretta, A. A. 2005. Reproductive ecology and behaviour of a species of Adenomera (Anura, Leptodactylinae) with endotrophic tadpoles: Systematic implications. Journal of Natural History 39:1745-1758. Fouquet, A., Ledoux, J. B., Dubut, V., Noonan, B. P., Scotti, I. 2012. The interplay of dispersal limitation, rivers and historical events shapes the genetic structure of an Amazonian frog. Biological Journal of the Linnean Society 106:356-373. Heyer, W. R. y Maxson, L. R. 1982. Distributions, relationships, and zoogeography of lowland frogs: The Leptodactylus complex in South America, with special reference to Amazonia. En Prance, T. P. (Ed.). 1982. Biological Diversification in the tropics. Columbia University Press: 375-388.PDF Lima, A. P. y Magnusson, W. E. 1998. Partitioning seasonal time: interactions among size, foraging activity and diet in leaf-litter frogs. Oecologia 116:259-266. Jetz, W., Pyron, R. A. 2018. The interplay of past diversification and evolutionary isolation with present imperilment across the amphibian tree of life. Nature Ecology & Evolution 2: 850-858.
|



 Map of Life
Map of Life