|
Vernacular Name
Cutín de Pichincha
State/Provinces
Esmeraldas, Pichincha, Santo Domingo de los Tsáchilas, Imbabura, Cañar, Guayas
Natural Regions
Bosque Húmedo Tropical del Chocó, Bosque Piemontano Occidental, Bosque Montano Occidental
Identification
Es una rana que va de muy pequeña a pequeña, dorso naranja pálido a café oscuro, vientre gris, ingles y superficies posteriores de los muslos amarillas. Algunos especímenes presentan cabeza anaranjada y el resto del cuerpo café, se puede observar una amplia franja dorsolateral anaranjada pálida. Presenta pequeños tubérculos en el talón. Los discos de sus dedos son ampliamente expandidos y carece de membrana basal entre los dedos de los pies. Es similar a Pristimantis walkeri, Pristimantis luteolateralis , Pristimantis esmeraldas y Pristimantis nietoi con quienes puede compartir su rango de distribución, pero se diferencia de las demás, porque las ingles y superficies ocultas de las extremidades tienen puntos de color amarillo anaranjado o amarillo limón sobre un fondo claro u oscuro. Pristimantis luteolateralis presenta un solo tubérculo grande en el talón y es de mayor tamaño (LRC machos = 22.0 mm y hembras = 27.9 mm; 13). Pristimantis esmeraldas y Pristimantis nietoi se diferencian por tener el vientre crema con varios niveles de café moteado. Además, Pristimantis esmeraldas nunca presenta los discos de sus dedos espadados (1, 3, 6, 10).
Description
Es una rana de tamaño variable entre muy pequeño y pequeño con la siguiente combinación de caracteres (3, 4): (1) piel dorsal levemente granular; vientre areolado; pliegue discoidal prominente; pliegues dorsolaterales ausentes; (2) cabeza ligeramente más ancha que el cuerpo; hocico corto, redondeado en vista dorsal y angularmente redondeado en vista lateral; fosas nasales protuberantes; canto rostral redondeado, pero evidente; región loreal cóncava; (3) membrana y anillo timpánico evidentes, redondos, ¼–⅖ del tamaño del ojo; (4) párpado superior sin tubérculos, usualmente más angosto que la distancia interorbital; crestas craneales ausentes; (5) odontóforos vomerinos ovales, no prominentes; (6) machos con hendiduras vocales; saco vocal subgular; almohadillas nupciales ausentes; (7) Dedo I de la mano más corto que el Dedo II; discos ampliamente expandidos; (8) dedos de las manos sin rebordes laterales; (9) tubérculos ulnares ausentes; (10) talón con tubérculos pequeños; (11) tubérculo metatarsal interno oval, cuatro veces el tamaño del externo redondeado; tubérculos supernumerarios en la base de los Dedos II y IV del pie; (12) dedos de los pies con rebordes laterales pequeños; membrana ausente; Dedo V del pie mucho más largo que el Dedo III.
Color in Life
Dorso varía de café claro a oscuro con tonalidades rojizas o grisáceas y puede tener marcas como líneas dorsolaterales, banda mediodorsal, barra interorbital o puntos oscuros. Barras labiales en la cabeza y línea supratimpánica. Tiene barras transversales oscuras en las extremidades. Las ingles y superficies ocultas de sus extremidades son amarillas anaranjadas a amarillas limón. Su vientre es gris a casi negro. El iris es café grisáceo a rojizo (3).
Males Face Cloacal Length Average
promedio = 18.0 mm (rango 15.5–19.6 mm; n=27) (3
Females Face Cloacal Length Average
promedio = 23.4 mm (rango 18.4–25.9 mm; n=11) (3
Habitat and Biology
Habita en bosque nublado y en zonas bajas tropicales de bosque lluvioso. Especie nocturna y arbórea, asociada a vegetación media sobre el suelo de borde de bosque (< 3 m). Durante el día, los individuos se pueden encontrar entre la hojarasca y en las axilas de la planta oreja de elefante. Sin embargo, McDiarmid encontró un individuo en una bromelia a 30 m de altura en un árbol, lo que indica que esta especie también se encuentra en el dosel del bosque ( 3, 4, 6).
Reproduction
Se han registrado hembras grávidas con 35 a 38 huevos maduros, en los meses de abril y agosto. Los machos vocalizan desde vegetación herbácea y arbustiva de sotobosque de bordes o interior de bosque primario y secundario, no en áreas abiertas (3, 6).
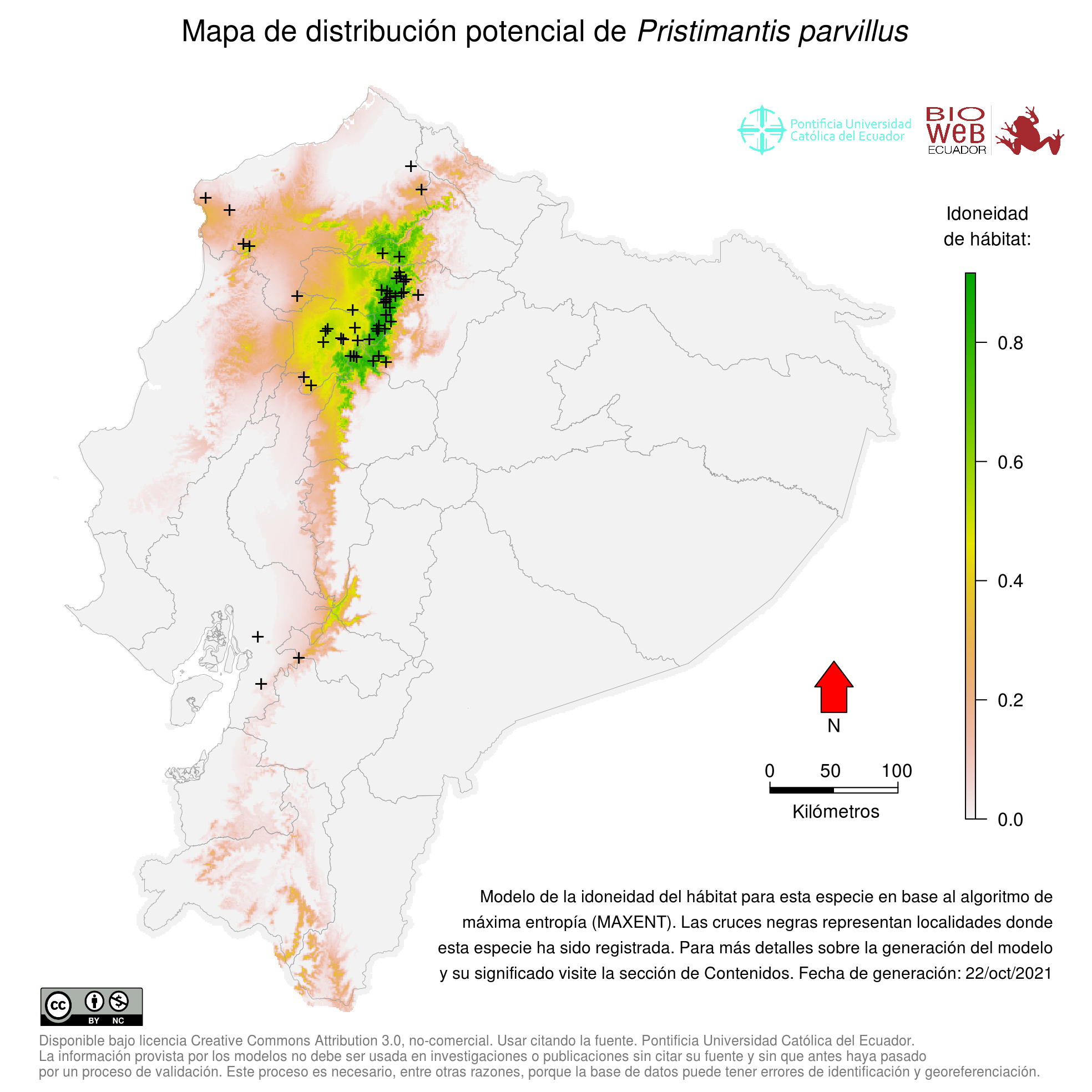
Global Distribution
Altitudinal Range
De 20 a 2000 metros sobre el nivel del mar
Taxonomy and Evolutionary Relationships
Está cercanamente relacionado a Pristimantis luteolateralis y Pristimantis walkeri ( 7 , 8, 9, 14). Asignado al grupo de especies Pristimantis unistrigatus ( 12). No ha sido asignado a ningún grupo de especies según Padial et al. ( 9).
Etymology
El epíteto específico es un diminutivo del latín parvus que significa diminuto, en referencia a que esta especie fue la más pequeña de las encontradas en Tandapi (4).



Summary Author
Caty Frenkel, Nadia Páez-Rosales, Andrea Varela-Jaramillo, Juan M. Guayasamin y Gabriela Pazmiño-Armijos
Editor
Mario Yánez-Muñoz
Edition Date
2016-06-07T00:00:00
Update Date
2022-09-28T21:42:39.387
Literature Cited
Guayasamin, J. M. 2004. A new species of Eleutherodactylus (Anura: Leptodactylidae) from the lowlands of northwestern Ecuador. Herpetologica 60:103-116.PDF Jiménez de la Espada, M. 1871. Faunae neotropicalis species quaedam nondum cognitae. Jornal de Sciencias, Mathematicas, Physicas e Naturaes. Academia Real das Sciencias de Lisboa 3:57-65. Lynch, J. D. y Duellman, W. E. 1997. Frogs of the genus Eleutherodactylus in Western Ecuador: systematics, ecology, and biogeography. The University of Kansas, Natural History Museum, Special Publication 23:1-236.Enlace Lynch, J. D. 1976. New species of frogs (Leptodactylidae: Eleutherodactylus) from the Pacific Versant of Ecuador. Occasional Papers of the Museum of Natural History, The University of Kansas, Lawrence, Kansas 55:1-33.PDF Kattan, G. H. 1984. Ranas del Valle del Cauca. Cespedesia 13:316-340.PDF MECN. 2010. Serie Herpetofauna del Ecuador: El Choco Esmeraldeño. Monografía. Museo Ecuatoriano de Ciencias Naturales. Quito-Ecuador 5:1-232. Pyron, R. A. y Wiens, J. J. 2011. A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Molecular Phylogenetics and Evolution 61:543-583. Pinto-Sánchez, N. R., Ibañez, R., Madriñan, S., Sanjur, O. I., Bermingham, E., Crawford, A. J. 2012. The great American biotic interchange in frogs: multiple and early colonization of Central America by the South American genus Pristimantis (Anura: Craugastoridae). Molecular Phylogenetics and Evolution 62:954-972. Padial, J. M., Grant, T., Frost, D. R. 2014. Molecular systematics of terraranas (Anura: Brachycephaloidea) with an assessment of the effects of alignment and optimality criteria. Zootaxa 3825:1-132. Arteaga, A. F., Pyron, R. A., Peñafiel, N., Romero-Barreto, P., Culebras, J., Bustamante, L. M., Yánez-Muñoz, M. H., Guayasamin, J. M. 2016. Comparative phylogeography reveals cryptic diversity and repeated patterns of cladogenesis for amphibians and reptiles in northwestern Ecuador. PLoS ONE 11(4): e0151746. doi:10.1371/journal.pone.0151746 Castro, F. y Cisneros-Heredia, D. F. 2004. Pristimantis parvillus. The IUCN Red List of Threatened Species. e.T56830A11541927. http://dx.doi.org/10.2305/IUCN.UK.2004.RLTS.T56830A11541927.en . Hedges, S. B., Duellman, W. E. y Heinicke, M. P. 2008. New World direct-developing frogs (Anura: Terrarana): Molecular phylogeny, classification, biogeography, and conservation. Zootaxa 1737:1-182. Ron, S. R. 2018. Base de datos de la colección de anfibios del Museo de Zoología (QCAZ). Versión 1.0. Pontificia Universidad Católica del Ecuador. Disponible en https://bioweb.bio/portal/ Jetz, W., Pyron, R. A. 2018. The interplay of past diversification and evolutionary isolation with present imperilment across the amphibian tree of life. Nature Ecology & Evolution 2: 850-858.
|


 Map of Life
Map of Life